Il y en a quelques unes sur la Vache notamment.
Rendez-vous sur la page colles. J’essaierai d’en mettre encore d’autres pendant ces vacances.
Répondre

Il y en a quelques unes sur la Vache notamment.
Rendez-vous sur la page colles. J’essaierai d’en mettre encore d’autres pendant ces vacances.
 Je participe aux ouvrages pour les BCPST chez l’éditeur Ellipses. J’ai relu une partie du livre de Biologie de 1ère année que vous pouvez commander par exemple ici. Vous pouvez même feuilleter une partie du premier chapitre là. J’aime beaucoup ce livre pour la simplicité des illustrations. Elles sont claires, synthétiques et facilement refaisables pour l’élève. Le texte est aussi tout à fait abordable et la mise en page particulièrement efficace
Je participe aux ouvrages pour les BCPST chez l’éditeur Ellipses. J’ai relu une partie du livre de Biologie de 1ère année que vous pouvez commander par exemple ici. Vous pouvez même feuilleter une partie du premier chapitre là. J’aime beaucoup ce livre pour la simplicité des illustrations. Elles sont claires, synthétiques et facilement refaisables pour l’élève. Le texte est aussi tout à fait abordable et la mise en page particulièrement efficace
J’ai écrit une partie du livre de Géologie qui ne devrait pas tarder à sortir. J’ai hâte de voir le résultat!
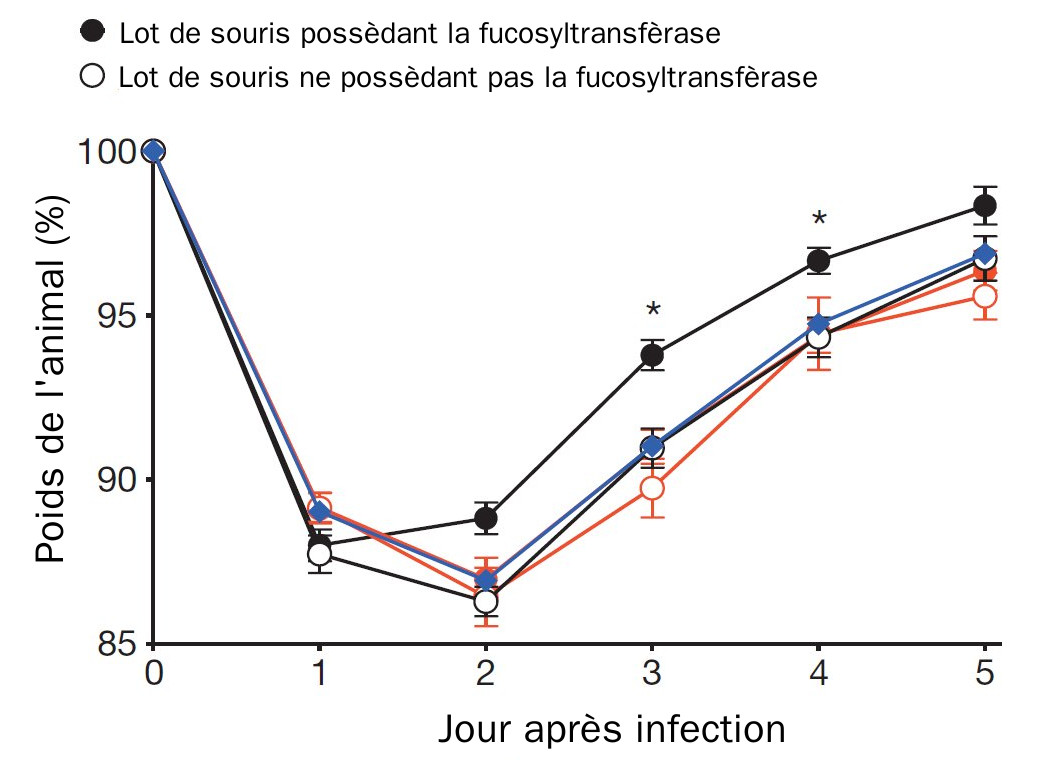
La réponse à une infection inclus la résistance au pathogène ou bien sa tolérance. Une infection est souvent associé à un anorexie temporaire, le malade se nourrit beaucoup moins. Ce mécanisme permet de relocaliser l’énergie provenant des aliments vers une résistance à l’infection mais permet aussi de priver le pathogène de nourriture. Cependant, cela aussi induit un stress sur la communauté microbienne constituant la flore intestinale puisqu’elle récupère, elle aussi, moins de nutriments. Ceci pose un problème à l’organisme puisque sa capacité de digestion s’en voit amoindrie d’une part et cela diminue la taille de la population de la flore intestinale qui protège elle aussi des pathogènes.
Dans un article paru cette semaine dans Nature, Pickard et ses collaborateurs mettent en évidence un mécanisme qui résout ce paradoxe. Lors de l’infection, des substances immunitaires, les inerleukines, sont produites en réaction aux pathogènes. L’équipe de Pickard montre ces interleukines entraînent aussi au niveau de l’épithélium intestinal la fucosylation, c’est à dire l’ajout de groupement fucose, un ose (correspondant à un galactose désoxydé au niveau du carbone 6) à certaines protéines par l’activation d’une fucosyl-transfèrase. Ces protéines sont ensuite exportés dans l’intestin ou le fucose est relâché par les protéines. Ceci a pour conséquence de diminuer l’expression des gènes de virulence des bactéries pathogènes ayant infecté le tube digestif mais permet aussi à la flore intestinale de récupérer des nutriments en utilisant le fucose ainsi produit et exporté par l’hôte. Ainsi, l’hôte se remet plus vite de l’infection et regagne plus rapidement son poids de départ.
Le rapport est paru et vous pouvez donc le trouver sur cette page
La domestication est un phénomène qui a le plus souvent eu lieu par sélection artificielle mais pas seulement. De plus, certains animaux sont plus facilement domesticables que d’autres.
Les résultats de l’admission sont publiés sur Publinet ce vendredi 4 juillet. Il se trouvent donc ici.
Félicitations à tous!
Les résultats de la session 2013 bis ou 2014 A sont là
Félicitations à tous!
J’en ai récupéré une partie…
Option A : Etude du protoplasme d’épinard, puis étude avec le DCPIP de la photosynthèse et enfin un exercice sur des mutants de la chaines photosynthétiques.
Option B : ESCARGOT : dissections (appareil digestif, système nerveux, appareil reproducteur), des documents sur la reproduction, prélever la radula et le dard, faire des tests pour déterminer leurs compositions. FUCUS : étude de l’appareil reproducteur, de l’étagement d’autres algues sur l’estran… FRUITS/GRAINES (mais, fusain…). Déterminations : associer des adultes (fourmi lions, roussette, mante religieuse etc…à la forme larvaire/oeuf/juvénile et décrire le mode de vie, de reproduction etc).
Option C
Contre-Option A :
Contre-Option B : La domestication du blé (étude comparative quantitative et qualitative de blés), étude du vol des criquets avec dissections et enfin reconnaissances de races de vaches et de végétaux/animaux
Contre-Option C : carte d’hydrogéologie, coupe, glacier à légender, comprendre l’origine des variations de concentrations en sulfate des eaux de sources en fonction des saisons titrage sur l’eau de Volvic pour calculer la concentration des ions en HCO3-