L’agrégation externe de SVT/SV-STU 2021 se déroule en ce moment. Voici les sujets :
Lire la suite
Répondre

L’agrégation externe de SVT/SV-STU 2021 se déroule en ce moment. Voici les sujets :
Lire la suiteLa datation par concordia-discordia est une technique assez utilisée en géoscience car elle permet de dater d’une part la mise en lace d’une roche magmatique mais aussi la réouverture du système isotopique, c’est à dire une modification des conditions à laquelle est soumise cette roche magmatique. La réouverture se faisant la plupart du temps par une augmentation de température, on date alors un événement métamorphique.
L’utilisation de base est simple puisqu’elle ne nécessite pas de calcul mais seulement une lecture graphique et sa compréhension.
Lire la suiteLes écrits de l’agrégation externe 2019 se déroulent du 11 au 13 mars. Voici les sujets qui sont tombés :
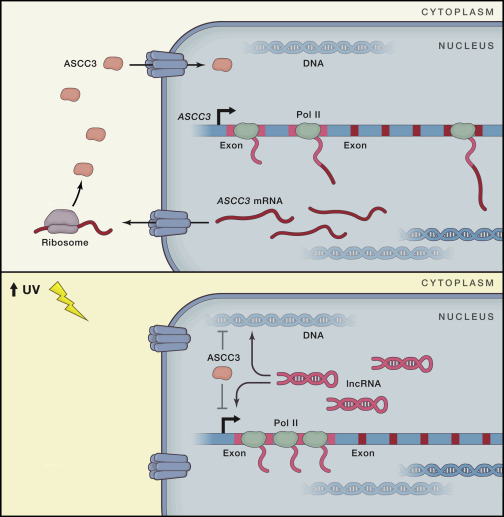
Lorsqu’une cellule est soumis aux UV, on sait que cela a comme conséquence d’entrainer des dommages à l’ADN. Une autre conséquence aussi est de diminuer la transcription. Dans un papier datant de février, Williamson et ses collègues découvrent l’implication d’une protéine et d’un ARNlnc dans l’arret et la reprise de la transcription après réparation. Là, où l’article est particulièrement interessant c’est que la protéine et l’ARNlnc proviennent du meme gène appellé ASCC3! Les travaux des chercheurs démontrent la chose suivante :
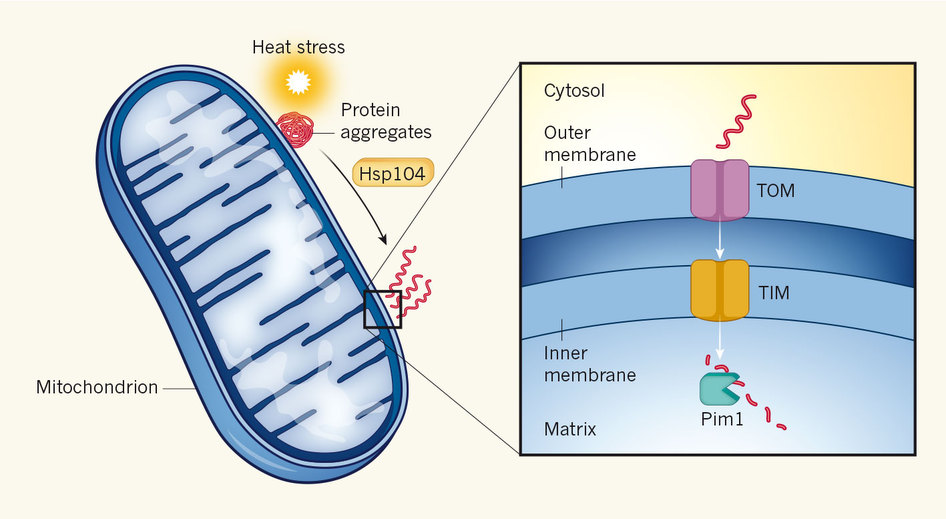
Lors d’un choc de température, on sait que la quantité de protéines mal reployées va augmenter : en effet, si la température augmente, l’agitation moléculaire augmente donc cela peut casser des liaisons faibles entre les radicaux des acides aminés, donc en mettre en place d’autres, donc indure un mauvais reploiement. On sait aussi que certaines protéines, les HSP, sont normalement chargées d’empêcher cela. Cependant, il y a tellement de protéines dans la cellule qu’une partie va forcément échapper à la surveillance. Une partie de ces protéines mal reployées vont avoir tendance à s’agréger et donc à former des complexes qui pourraient etre dangereux. Ces complexes doivent donc etre détruits. Une structure impliquée dans la destruction des protéines est le protéasome mais ce n’est pas de lui dont il est question ici. Dans un article publié dans Nature, Ruan rapporte que dans des conditions de stress thermique, les agrégats de protéines formées ont tendance à s’accumuler au niveau de la surface des mitochondries. Une protéine chaperon, HSP104, désagrège l’agrégat, et une partie est ensuite transportée à l’intérieur de la mitochondrie via le complexe TOM-TIM. Une fois dans la mitochondrie, la protéase Pim1 dégrade les protéines qui sont entrées si elle ne sont pas destinés au fonctionnement de la mitochondrie.
Bose revient sur le role des séquences cis-regulatrices et ajoute un nouveau type d’ARN et une nouvelle entorse au mécanisme canonique de transcription. En effet, dans Cell, Bose et ses collègues montre l’importance d’un nouveau type d’ARN les eRNA. Ces eRNA sont des produits provenant de la transcription de séquences cis-régulatrices (eRNA veut dire ARN enhancer donc transcrits à partir de séquences enhancer) et sont donc des ARN non-codants (quand je vous disais qu’on se rend compte en réalité que tout le genome à peu près est transcrit…). Au niveau de la chromatine, certaines protéines comme CPB sont chargés d’acétyler les histones mais sont inactives schéma a). Cependant, lors de la transcription des eRNA, alors qu’ils n’ont pas été encore totalement transcrits, ils vont pouvoir intéragir avec d’autres facteurs de transcription et CPB : cette intéraction ARN-protéine va activer CPB et lui permettre d’acétyler les histones ainsi que certains facteurs de transcription (schéma b). Cette acétylation entraine le recrutement de co-activateur de la transcription qui va permettre d’activer l’activité de l’ARN Pol II qui se met alors aussi à transcrire le gène d’interet (schéma c).

Le problème de l’initiation de la tectonique des plaques ainsi que de la mise en place de la subduction reste problématique voire énigmatique. En effet, à l’heure actuelle, l’initiation d’une subduction nécessite au préalable des forces aux limites et donc une tectoniquue des plaques ainsi que des zones de fragilité dans la lithosphère conséquence là encore de la tectonique des plaques.
Dans un papier publié dans Nature en décembre 2015, Taras GERYA propose une simulation numérique très interessante qui est en lien d’une part avec notre programme mais aussi qui permet de revenir sur une question à laquelle nous n’avons pas répondu et qui corrobore son hypothèse à savoir l’ouverture de l’océan Atlantique.
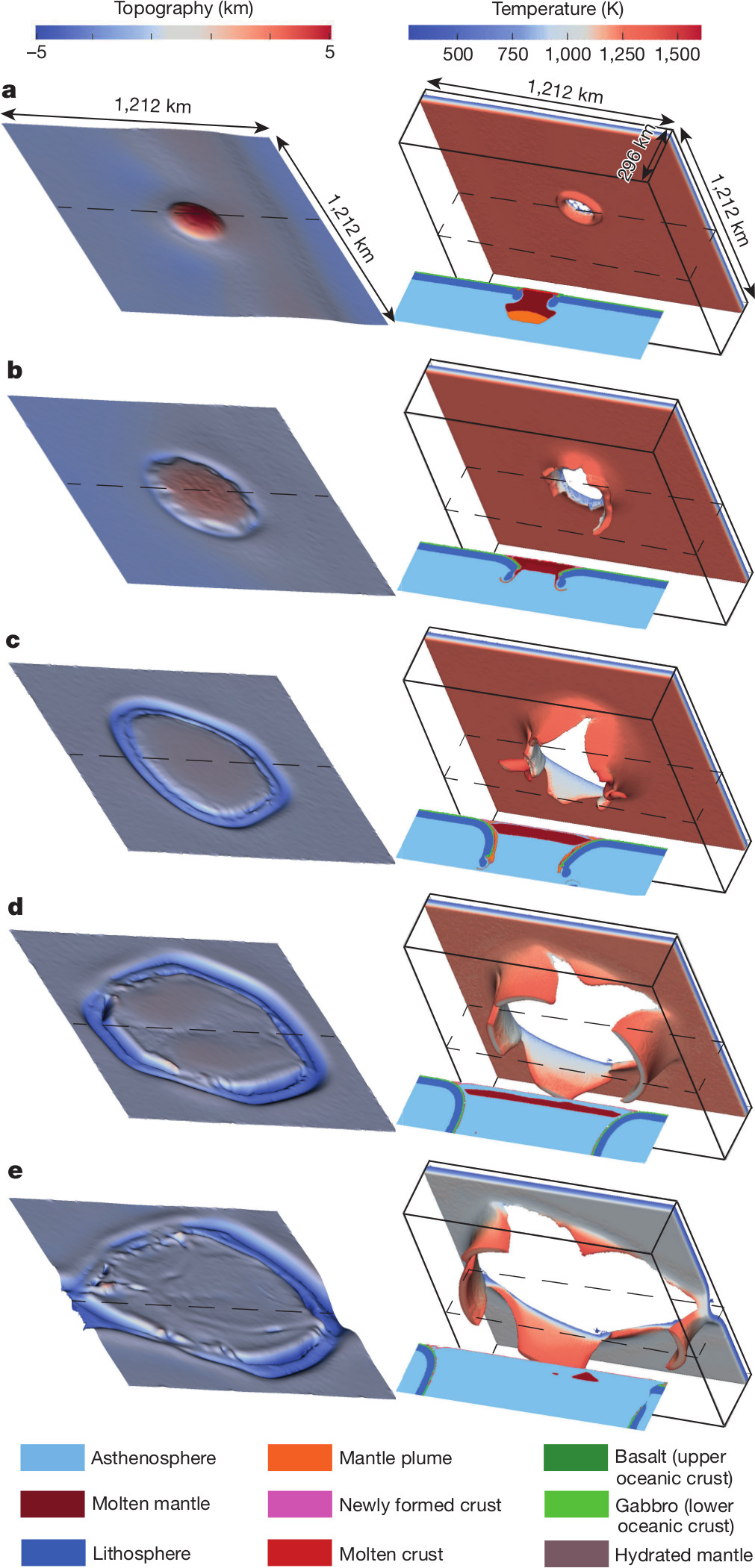 La simulation numérique ci-dessous montre la conséquence de l’arrivée d’un point chaud sur une plaque complètement homogène. Cette arrivée entraine la formation
La simulation numérique ci-dessous montre la conséquence de l’arrivée d’un point chaud sur une plaque complètement homogène. Cette arrivée entraine la formation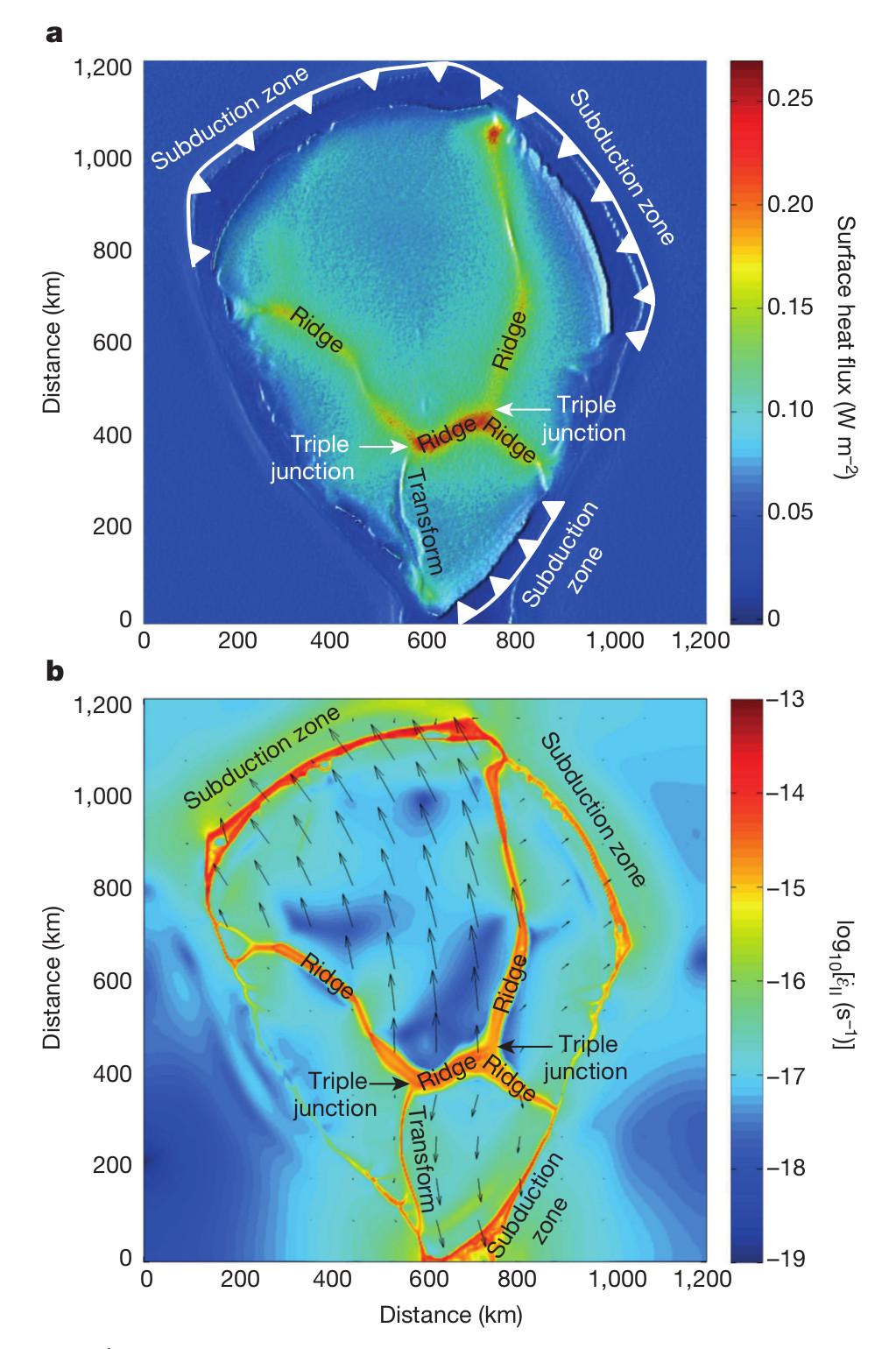
a) de ce qu’on appelle un plateau océanique puis
b) la formation d’une fosse et d’un slab (= un panneau plongeant de lithosphère dans le manteau) de forme circulaire puis
c) un déchirement du slab puis
d) la formation d’une véritable zone de subduction avec un retrait en arrière du slab (on en reparlera dans le cours sur le magmatisme) puis
e) la formation de dorsale et de faille transformante
La colonne de gauche montre la topographie , celle de droite la morphologie de la lithosphère subduite avec la projection de la température de surface du slab
Lorsqu’on regarde la surface de la Terre à partir de cette arrivée de point chaud comme montré à droite, on constate en effet la formation de plaque lithosphérique (a) animée de mouvement vers les zones de subductions (b)
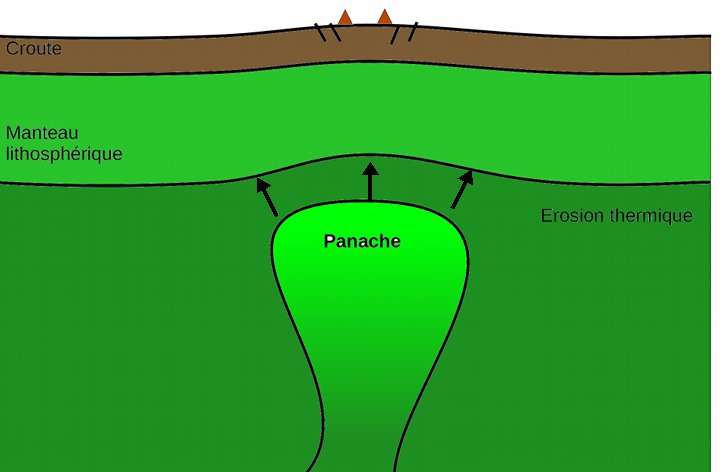
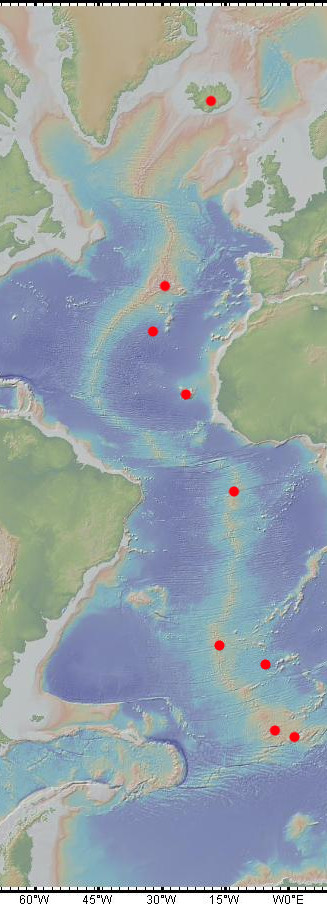 Non seulement cette simulation explique comment la tectonique des plaques a pu être initiée mais vient en appui à la théorie selon laquelle l’océan Atlantique aurait été ouvert par l’arrivée de point chaud. En effet, si on regarde la localisation des points chauds dans l’océan Atlantique, on constate qu’ils sont sur la dorsale ou très proche de la dorsale. Ces panaches mantelliques, en arrivant en surface entraine un réchauffement de la base de la lithosphère. Or, la base de la lithosphère est définie par l’isotherme 1300°C. Si la base est réchauffée, alors, elle remonte et la lithosphère devient donc plus fine à sa base. Par compensation isostasique, la partie supérieure de la lithosphère s’effondre ce qui entraine un rifting : c’est le rifting actif.
Non seulement cette simulation explique comment la tectonique des plaques a pu être initiée mais vient en appui à la théorie selon laquelle l’océan Atlantique aurait été ouvert par l’arrivée de point chaud. En effet, si on regarde la localisation des points chauds dans l’océan Atlantique, on constate qu’ils sont sur la dorsale ou très proche de la dorsale. Ces panaches mantelliques, en arrivant en surface entraine un réchauffement de la base de la lithosphère. Or, la base de la lithosphère est définie par l’isotherme 1300°C. Si la base est réchauffée, alors, elle remonte et la lithosphère devient donc plus fine à sa base. Par compensation isostasique, la partie supérieure de la lithosphère s’effondre ce qui entraine un rifting : c’est le rifting actif.
(Le programme de la session 2014 est reconduit pour la session 2015)
Le programme du concours inclut l’ensemble des programmes des classes de collèges et de lycées que le futur enseignant de sciences de la vie et de la Terre devra maîtriser et huit thématiques plus spécialisées.
Le niveau de maitrise de ces thématiques est un niveau universitaire permettant d’avoir le recul attendu d’un enseignant disciplinaire, tant sur les connaissances, que sur les méthodes ou les démarches.
Liste des 8 thématiques spécialisées :